El Aparato de Golgi
El Aparato de Golgi es un orgánulo de las células eucariotas. Puedes ver más acerca de los orgánulos visitando nuestro artículo “orgánulos: ordenando la célula” aquí.
El aparato de Golgi fue descrito en 1876 por el médico italiano Camillo Golgi mientras estudiaba la estructura del tejido nervioso. En 1906 obtendría el premio Nobel de Medicina, por sus estudios en la células que conformaban el sistema nervioso. Compartió el premio con Santiago Ramón y Cajal, que basándose en la tinción desarrollada por Golgi enunció la teoría neuronal.
Localización celular: el Golgi es un orgánulo citoplasmático que se sitúa entre el Retículo endoplasmático (RE) y la membrana celular (puedes saber más del retículo endoplasmático aquí). Mantiene una estrecha relación con el RE y éste a su vez con el núcleo, por lo que forman la vía común de tránsito entre el núcleo y la membrana para el transporte de proteínas desde su síntesis en el núcleo hasta su destino.
Descripción citológica de su estructura: El Aparato de Golgi, también llamado Dictiosoma, está formado por varias cisternas o sáculos formados por membrana, dependiendo de la célula pueden tener entre 4 y 80 sacos. Estas cisternas tienen una forma aplanada, con su cara convexa hacia el RE. Se colocan en sucesión desde el RE hacia la membrana. Estas cisternas están comunicadas entre ellas por tubos membranosos y por las vesículas membranosas que intercambian.
Dependiendo de su localización podemos establecer tres zonas en el AG. La región del cis-Golgi es la más cercana al RE y su composición lipídica y de proteínas es similar al RE. La región medial, situada a continuación del cis-Golgi comunica ésta región con la siguiente, su composición molecular es un punto intermedio entre las dos regiones adyacentes. El trans-Golgi esta región es la más cercana a la membrana plasmática y su composición es similar a membrana citosólica.
Además de las tres regiones forman parte del Golgi todas las vesículas que están viajando entre dos de sus cisternas.
Función: en el AG se modifican tanto proteínas como lípidos sintetizados en el RE, liso y rugoso, ambos tipos de moléculas viajan en las vesículas que salen del RE y se unen a las cisternas membranosas del AG, las cisternas contienen enzimas necesarios para la modificación de ambas moléculas, tales como agregación de grupos fosfato o carbohidratos (fosforilación y glicosilación, respectivamente). Tales añadidos entran en las cisternas del AG desde el citoplasma y a medida que las proteínas o los lípidos viajan a través del AG, entrando y saliendo de las cisternas en vesículas van adquiriendo tanto los grupos que les faltan como del mismo modo marcadores que ayudarán a señalizar el destino de la molécula. Por ejemplo la manosa-6- fosfato es un marcador para que la molécula viaje al lisosoma.
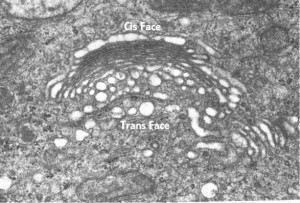
Al microscopio electrónico puede verse las cisternas y vesículas del AG.
Una vez alcanzada la región trans del AG los lípidos forman vesículas con las proteínas y viajarán por el citoplasma hasta su destino, que en ocasiones es el exterior celular, normalmente la matriz celular. Al unirse las vesículas a un orgánulo o a la membrana celular la membrana no solo recibe las proteínas que contuviese la vesícula sino que también aumenta de tamaño por los lípidos que formaban la vesícula. Por esto las vesículas que salen del Golgi pueden ir tanto a la membrana como “hacia atrás”, a la membrana del RE o a la del núcleo.