La bioquímica de la Acetilcolina
La acetilcolina es un neurotransmisor que se produce en el terminal nervioso a partir de acetil-CoA producido en las mitocondrias a partir de glucosa y la colina. No está claro cómo sale el acetil-CoA al citoplasma, pero allí la acetilcolina sintetasa o colina acetiltransferasa (ChAT) produce finalmente la acetilcolina. Una vez sintetizada se lleva a vesículas de almacenamiento mediante gasto de ATP.
La colina para la formación de la acetilcolina puede ser adquirida en la dieta o por recaptura del espacio intercelular en la misma neurona que la usa como neurotransmisor. Esta recaptura permite un gran ahorro energético al evitar tener que sintetizar colina de novo. La afinidad de la enzima por el acetil-CoA es muy grande, por lo que se cree que la limitación de los depósitos de acetilcolina depende principalmente de las cantidades de colina disponibles.
Internalización de la colina
La colina entra a las neuronas colinérgicas por un cotransportador de alta afinidad dependiente de Na+ y Cl- (ChT). La concentración de colina es de 10 a 100 ?mol/L, y estos transportadores suelen tener una KM de 1-5 ?mol/L. Con estas concentraciones el transportador se satura, y así se limita la entrada de colina.
También existe un transportador de baja afinidad con KM de 10-100 ?mol/L, que no es saturable. No depende de Na+ y que se halla presente en todos los tejidos.
Relleno y liberación de las vesículas sinápticas con acetilcolina
Desde el citosol la acetilcolina debe pasar a la vesícula sináptica Para tal fin existen unas proteínas transportadoras de vesículas de acetilcolina (VAChT), con características similares a otros transportadores vesiculares. Su estructura consta de 12 hélices de transmembrana, y un N-ter y C-ter en posición extracelular. Llevan acetilcolina al interior con un cotransporte antiporte con 2H+. El gradiente de pH necesario se consigue mediante la acción de una ATPasa que bombea H+ al interior contra gradiente aprovechando la energía de hidrólisis del ATP. La vesícula es ácida pero electroneutra, ya que en el interior hay un polisacárido (vesiculina) con carga negativa que neutraliza las cargas. La vesícula puede contener unas 5 000 moléculas de neurotransmisor, unas 50-100 mM.
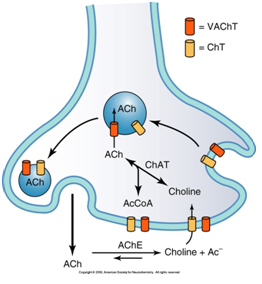
Esquema de la síntesis y la recaptura de la Acetilcolina en el botón sináptico.
Hay 2 tipos de vesículas de acetilcolina. Unas son pequeñas y densas, y contienen el neurotransmisor recién formado. Cuando llega un potencial de acción a la neurona son las primeras en liberarse. Además están muy cercanas a la membrana neuronal. Otras vesículas son más grandes y menos densas, y ante la llegada del potencial de acción no se liberan al estar más alejadas de la membrana. Son vesículas de reserva que suministran acetilcolina al pool de acetilcolina de rápida liberación.
Se libera acetilcolina, vesiculina y moléculas de ATP, ya que se vacía todo el contenido vesicular. Lo interesante es la acetilcolina, que se une a receptores específicos, y después se degrada en acetato y colina. La colina puede metabolizarse a betaína y después a serina, o se puede recapturar.
La acetilcolinesterasa degrada la acetilcolina en acetato y colina. Tiene una estructura globular catalítica, que forman monómeros, dímeros o tetrámeros según los diferentes enlaces químicos que se establezcan:
• Monómeros: el grupo tiol permanece reducido
• Dímeros: dos subunidades se unen por un puente disulfuro
• Tetrámeros: dos dímeros se unen mediante fuerzas de van der Waals.
Hay una forma adicional, que se produce cuando un tetrámero se asocia a una cola de colágeno mediante dos puentes disulfuro, y se asocian tres de ellas por enlaces iónicos para dar lugar a 3 tetrámeros asociados a una triple hélice de colágeno, a modo de ramillete. En esta composición se adquiere la capacidad de anclarse a membrana en la hendidura sináptica.
Hay una secuencia de aminoácidos importante para la catálisis en la acetilcolinesterasa. La enzima tiene cargas – gracias al Glu 327 para atraer a la acetilcolina con carga +. Además hay un grupo –OH de una serina que representa un punto esterático, y que le permite interaccionar con el grupo carbocarbonilo de la acetilcolina. Se forma un complejo que luego se va a romper, y deja libre la colina y la molécula acetilada. Una molécula de H2O facilita la pérdida del acetilo.